Research Interests
SPIDERS WE LIKE SPIDERS

Evolutionary origins of high performance major ampullate spider silk
Spiders are among the most diverse animals on earth, due in part to the exceptional properties of their silk. These properties are epitomized by orb weaving spiders that spin the backbones of their webs using dragline silk, which has a high tensile strength and extreme toughness, exceeding Kevlar®. Thus, orb weaving dragline silk is an important model for materials science, but it is an ancient silk and many other spider lineages also spin dragline silk. This project seeks to understand how dragline silk evolved such astonishing characteristics by focusing on its evolutionary change at the genetic, biomechanical and ecological levels. Objective 1 compares the structure and function of dragline silk from ancient lineages of spiders to tarantulas, which spin a primitive type of silk. Objective 2, takes supercontraction, a novel property of dragline silk that may allow spiders to control fiber spinning, and tests key hypotheses about when it evolved and what causes it. Finally, Objective 3 tests the role of ecological changes in dragline use as a force shaping the evolution of dragline’s material properties. This project will yield a better understanding of a biomaterial critical to the success of a mega-diverse lineage of animals and also provide valuable insight for the synthesis of biomimetic dragline silk analogs. The project will also play a key role in the development of the University of Akron’s brand new PhD program in Integrated Bioscience (IB), which is designed to train students in collaborative research at the interface between biology and other disciplines, by directly supporting and training students and by serving as a “hands on” case study of integrative research that will be followed by the first cohorts of these IB students.

Spinning physiology, supercontraction & silk muscles
The performance phenotypes of different silks are determined by a complex interplay between the amino acid sequences of silk proteins and how those proteins are assembled, or spun, into solid fibers. Water plays an essential role in the processing of silk fibers, which begin as liquid solutions, by maintaining mobility of protein secondary structures as fibers are spun. This likely provides spiders some degree of control over the molecular alignment of silk threads and therefore also the performance of silk and webs. It also results in a spectacular phenomenon called supercontraction where silk threads can shrink up to 50% of their length, generating substantial force. We are currently investigating the causes and consequences of supercontraction across spider taxa that are diverse in their ecological uses of silk and their silk production physiologies.
We recently discovered that spider silk displays a second, totally discrete, interaction with water where it expands and contracts repeatedly in response to changing humidity. In contrast to supercontraction, this cyclic contraction involves water induced relaxation, is highly repeatable, and scales precisely with the magnitude of changing humidity. Yet, it generates comparable forces. All of these qualities make spider silk a unique system for the development of a new type of biomimetic muscle, driven by changes in water content alone. Thus, spider silk may emerge as an important actuators for micro-robotics, drug delivery, sensor technology, or even green energy production. This work is in collaboration with Ingi Agnarsson and Ali Dhinojwala and is supported by NSF awards "Acquisition of a dynamic nano-force tensile test system for ultrathin fibers with environmental control and integrated image analysis" and "CAREER: Evolutionary origins of high performance major ampullate spider silk: development of an integrated bioscience training program".

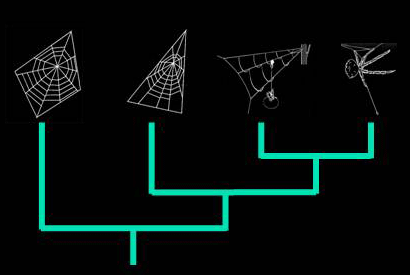
Reconstructing spider web evolution
A robust understanding of the evolutionary relationships among spiders is central to investigating relationships between web function and silk evolution. I am developing new phylogenetic hypotheses for how orb weaving spiders evolved and for evolutionary relationships among orb spiders. These phylogenies will then be used to hypotheses about coevolutionary relationships between silk function and structure, as well as the ecological determinants of spider diversification. This work emphasizes a combined approach utilizing morphological, behavioral and molecular data. This research is part of two NSF funded projects: "Araneid phylogeny and evolution of spider silk phenotypes" and "Acquisition of a dynamic nano-force tensile test system for ultrathin fibers with environmental control and integrated image analysis" in collaboration with Ingi Agnarsson, Jonathan Coddington, Volker Framenau, Cheryl Hayashi, Nikolaj Scharff and John Wenzel.

Diversification of Spider Silk Biomechanics
Orb weaving spiders spin silks from seven different glands, all of which are used for different functions. Some of these silks are as strong as steel, while others are as stretchy as rubber. These silks are constructed from a small array of proteins, providing unique opportunities to link external selective pressures on the mechanical performance of spider silks to evolutionary changes in silk genes.
Based upon research that I began in the laboratory of Cheryl Hayashi, I am currently characterizing the biomechanical properties of this diverse tool kit of silks for many spiders to study how the mechanical performance of silks changes during behavioral shifts in how spiders construct webs, such as the evolution of capture webs and the development of aerial orb webs (Blackledge et al. 2005a, b). Furthermore, I am studying how spiders can manipulate the mechanical properties of silk in behavioral contexts such as responding to different types of prey or constructing different types of webs. This research is part of two NSF funded projects: "Araneid phylogeny and evolution of spider silk phenotypes" and "Acquisition of a dynamic nano-force tensile test system for ultrathin fibers with environmental control and integrated image analysis" in collaboration with Nikolaj Scharff and John Wenzel.
Evolution of prey specialization in spiders
The Araneidae is the largest family of spiders that construct wheel-shaped two-dimensional orb webs. Yet, many araneids have evolved highly specialized web construction behaviors that result in webs unlike typical orb webs. I am currently working to establish a generic level phylogeny for relationships within this group to investigate how and why these behavioral specializations have evolved, how they contribute to the dominance of araneid spiders as predators of flying insects, and how the evolution of behavior and silk biomechanics interact with one another. This project includes molecular data from multiple nuclear and mitochondrial loci, as well as morphological data in collaboration with Dr. Nikolaj Scharff and Mark Stowe. The extreme web reduction found in bolas spiders is of particular interest to me. Bolas spiders have reduced the orb web to a single sticky thread, which they throw at moth prey. These moths are attracted through aggressive chemical mimicry of female moth sex pheromones. Our phylogenetic research is currently testing the hypothesis that this unique hunting behavior evolved through a series of web reductions in a number of closely related species of spiders that display lesser degrees of specialization upon moths. Once we have established an historical framework for the evolution of web building behaviors within these moth specialists, we will then examine changes in the functional properties of the unique capture silks used by these spiders. This will provide insight into how evolution has acted upon the genes coding for silks during extreme behavioral specialization.


Tetragnatha hawaiensis web from Hawai'i

Tetragnatha stelarobusta from Maui

Cyclosa simplicicauda rufescens from Hawai'i
Behavioral innovation and biodiversity: adaptive radiations in Hawai'i
Adaptive radiation is a major theme in the evolution of the world’s biodiversity, yet in non-adaptive radiations, speciation occurs with minor change in the ecologies of species. Little is known about whether lineages in adaptive radiations exploit more resources than non-adaptive radiations, divide resources more finely, or both? Furthermore, does adaptive radiation result in predictable evolutionary patterns of resource exploitation, or is historical contingency so great that, despite selective pressures in common between communities, evolution of resource use within radiations is largely unpredictable? I have found that within an endemic radiation of Hawaiian spiders, sympatric species of Tetragnatha display an extraordinary diversification in how and where they construct webs (Blackledge, Binford & Gillespie 2003). Remarkably similar web building behaviors, or “ethotypes”, have evolved independently in groups of species on different islands, suggesting a deterministic pattern to the behavioral diversification of endemic Hawaiian Tetragnatha (Blackledge & Gillespie, 2004). Future research will examine how these web ethotypes function in prey capture, to test whether convergence in web architecture is related to convergent selective pressures for prey capture behavior (e.g. Blackledge, Hang & Gillespie, in prep). Furthermore, I will compare the biomechanical properties of the capture silks used to construct these diverse webs, as a framework to study how silk gene evolution is related to behavioral diversification within this adaptive radiation.
Cylcosa (Araneidae) is the only other group of orb-weaving spiders, besides Tetragnatha (Tetragnathidae), to have dispersed to and speciated across the isolated Hawaiian Islands. In contrast to Tetragnatha, Cyclosa comprises a relatively species poor radiation. These two groups of spiders provide a unique opportunity to study why speciation is higher in one lineage than in another. Both genera are sympatric, found within the same habitats, but are segregated temporally because Cyclosa is diurnal while Tetragnatha is nocturnal. Thus, competition does not explain this difference in diversification. Preliminary data also suggest that the Hawaiian Cyclosa are not especially younger than Hawaiian Tetragnatha. Instead, comparison of resource use between these two radiations may give insight into the causes of adaptive radiation. By quantifying variation in the web architectures and resource use of endemic Hawaiian spiders and reconstructing their phylogenetic relationships I can answer two hypotheses about diversification. Do species diversify in adaptive radiations because they exploit greater ecological opportunity? Or, do species divide resources more narrowly, or tolerate greater niche overlap, so that adaptive radiations can occur regardless of ecological opportunity?

Argiope aurantia is common around Akron

immature Argiope often construct doily-like designs

mud-dauber wasps are important predators of spiders
Predator-prey conflict and sensory drive: stabilimentum web decorations
My research focuses on the conflict that arises, between signaling presence of webs to predators and to prey, when spiders include stabilimenta in their webs. These conspicuous silk lines, crosses and spirals may have several defensive functions including camouflage of spiders, startling predators, and acting as aposematic warnings for the presence of webs (Blackledge & Wenzel 1999, 2001). However, my research indicates that insect prey can also use stabilimenta as a signal in avoidance of webs, indicating that there should be selection against the use of stabilimenta in web avoidance by insects (Blackledge & Wenzel 1999). The reflectance spectrum of the silk used to build stabilimenta suggests that the silk is cryptic to insects, unlike more primitive spider silks (Blackledge 1998b). This is supported by my experiments demonstrating that honey bees can learn to forage at targets made from primitive spider silks but not targets made from stabilimentum silk (Blackledge & Wenzel 2000). I suggest that the evolution of silk coloration has occurred through a process termed sensory drive, where innate biases in the color vision of insects has selected for the cryptic properties of stabilimenta. This system is unusual because most examples of sensory drive involve sexually selected signals but spiders’ silks have evolved under natural selection from predators and prey.